

|
Secrets of spines Stereotype belief concerning the functions of cacti spines as the organs providing protection against sunrays and animals certainly does not cover the subject fully. Only detailed observation of spines through a microscope can reveal a lot of interesting biological puzzles. Most of them can be explained by the evolution process, whose role is to adjust plants as well as possible to the environment in which they make their living. Not all, however, as there are also some structures the function of which still remains a secret hidden in the area not accessible for the human eye. Basically every group of cacti has produced, throughout the evolution process, its own spine structure characteristics; very often significant differences occur for two cacti species of the same genus. The photographs presented in the article show spine built characteristic for the Pterocactus genus and for related groups belonging to sub-family of Opuntioideae. Internal structure of spines is identical for all nine (9) species of this genus. In their cross-section, it is possible to notice tiny plates joined together in quite a chaotic manner, and so creating a sponge-like structure. This is very light, due to empty spaces left by the plates but at the same time very resistant to mechanical forces. In this simple way, spines which do not require any strong points of attachment in areoles, are able to maintain their defensive functions.
|
| | |
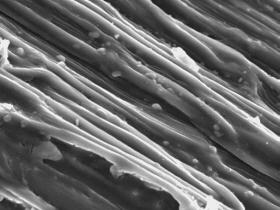
| General
view of a spine surface (P.fischeri, KG 2001, Vaca Muerta) |
Fibres on a spine surface
|
Cross-section, outer tunicle
and inner structures of a spine |
|
The structure described above can also play a part in two further factors: First, sunrays do not heat spines to such extent as they would do for homogenous structure. Secondly, rain
water and condensed steam can penetrate the free spaces within freely, gather
Putting spines into water for some time can easily check the phenomenon described. The effect of liquid absorption results in spines becoming more flexible and their losing defensive properties, then practically not pricking the skin. The outer layer of spines is built in a similar way. Plates there, being considerably bigger, are layered horizontally around the core making a coating even more resistant to weather factors' operation. Joining together, they sometimes tend to make up longer channels filled with air, but generally they remain the layer tightly covering the internal part of the spine. The most outer layer is at the same time the most interesting one, as it often creates interesting forms. Seeing it under the microscope lens it is hard to discern its constituting parts. Differently than in the inner structure, here appear characteristic fibres intertwined with each other and giving the whole layer proper resistance to mechanical forces which may be applied to it. The weave of those fibres could be compared to a rope structure, which is much more durable than a single fibre of the same cross-section. At the ends of spines, most fibres bend towards the outside, creating specific appendixes, or acanthas. Their mode of functioning is easy to guess. Spine goes into the skin easily, but it is much more difficult to take it out, since the appendixes work similarly to fishing hooks. One may wonder why in its evolution process the plant has created a similar mechanism which encourages breaking off consecutive sprouts. In my opinion, some explanation would be the hypothesis assuming this way to be plant's vegetative reproduction in natural habitat. In case of Pterocacti this seems fairly probable, as two of its species, P. kuntzei and P. megliolii, do have the ability to lose their sprouts in autumn and winter. These are then spread by strong winds and volatile sands, and can give origins to a new plant in the next vegetative season. The remaining species do not have this property, however long drought also causes easy breakage of sprouts. Unfortunately, confirming or abolishing this particular, reproductive function of spine hooks still remains within theoretical considerations. I would like to point out this opinion to be my own only, not supported by any observation in natural habitat of cacti, which are the essential part of the hypothesis in this case. Another possibility cannot be excluded: perhaps we are witnessing a constant evolution process striving to create such reproductive mechanism. It should also be noticed that similar hooks on glochides are to set in the intruder's skin and cause longer discomfort. As opposite to main spines, however, glochides have very little attachment within areoles, and so can be easily removed from them.
|
 |  |  |
|
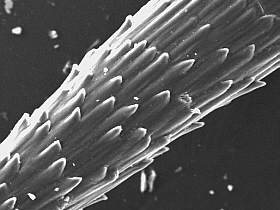
Inner structures of
a spine, structure tiles (P.hickenii, KG 2466, Perito Moreno) |
Specific hooks on a spine
surface |
. Close - up of specific
hooks |
|
Similarly, attention should be drawn to a phenomenon common in animal world, namely sacrifying some part of the individuals or some part of animal itself in case of predator's attack. Examples of such defence mechanisms providing survival can be the strategy of self-defence presented by bees. Stinging an intruder, they also do let go some of their inner organs, therefore being sentenced to die in defence of other individuals, and most of all in defence of the queen. Many species of starfish act the same way; in the moment of danger their main shield may spontaneously be separated from some of their arms. Both parts will have high potentials to regenerate the lost parts of the body, but only one will have a chance to survive the other becoming the attacker's prey. This specific property is also used for reproduction purposes. Basing on the above examples it can be concluded that the function of hooks on ends of the main spines may be limited only to plant's protection from predators and have no role in vegetative reproduction. It should also be noticed that both functions mentioned above could be complementary. We have to always remember, that evolution is caused by totally accidental mutations, though due to existance of natural selection, it may be expected, that some profitable feautures will be preserved. Taking photographs discussed in the article under an electron microscope, my hopes were also to characterise the spine surface of further Pterocactus species. Among other things, my aim was to prove or to reject the thesis of skottsbergii form belonging to the P. hickenii species. The limits between other types of this species are not clear-cut either. Some specialists point out existence of intermediate forms between such species as P. australis and P. fischeri and between P. hickenii and P. araucanus (Klaus Gilmer). Unfortunately it turns out that mere characteristics of spine surface cannot provide unambiguous answers to these questions. Seen through an electron microscope, structures occurring in a some millimetres long spine P. gonjianii are the same as in a much bigger spine of P. hickenii. Nevertheless, in P. australis I was able to notice a separate spine surface structure reminding, in simplified comparison, bark of coniferous trees. In my opinion, P. australis may be a separate species, a one not having direct correlate in spite of its high degree of variability. Even in this case, however, too hasty conclusions may prove harmful. It is certainly necessary to do more, and more detailed, in this subject and to compare spine surfaces of a bigger number of separate clones. |
 |  |  | |||
|
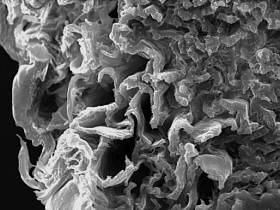
Cross - section, outer
tunicle and inner structure of a spine
|
Cross - section, specific
tubules in inner
|
Specific hooks on end-piece
of a spine
|
|
As far as chemical composition of spines is concerned, there could not have been expected any surprises. As in every biological specimen, in spines there occur almost only carbon and oxygen compounds. Chemical elements of values below 10 degrees fall into the background.
|
 |  |  | |||
|
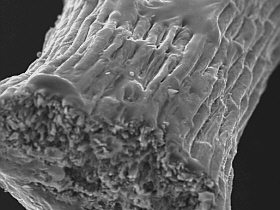
Fibres on a spine surface | Surface
of a spine and a delicate hair |
General view of inner
and outer structures |
 | |||||
|
Surface of a spine
| ||
| Please contact me for Critical and let me know your opinion. E-mail-Piotr Swiatoniowski Special thanks
for Klaus Gilmer, who kindly sent the cuttings, which render this article possible
to write. | ||||